Database Building
Sequences
The database is built using sequences taken from SWISS-PROT and its annex TrEMBL. The reasons that led us to choose these collections are threefold:
- The SWISS-PROT + TrEMBL set is exhaustive and non-redundant.
- The annotations are of high quality compared to the other general sequence database systems.
- Almost all entries are cross-referenced with their corresponding nucleotide sequences in GenBank / EMBL / DDBJ.
Duplication
In SWISS-PROT+TrEMBL, homologous genes from different organisms (or paralogous genes within a
genome) that code for a same protein are described by a single entry.
In the view of HOVERGEN, there should be a specific entry for each gene to allow one to retrieve all
existing homologous genes. For these reasons the SWISS-PROT + TrEMBL entries that correspond to
several genes are duplicated before being used for families calculation. These duplicated entries are
given a new name. The duplicated sequences are signaled by their annotations in the CC field:
Example: The sequence 143G_MOUSE3 presents the
following annotation:
CC -!- modified from 143G_HUMAN.
Families
To build the families we perform a similarity search of all the proteins against each other with BLASTP2. For this purpose, we use the BLOSUM62 similarity matrix and a threshold of 10-4 for E-values. Low complexity sequences are filtered with SEG. Then, the results are processed this way:
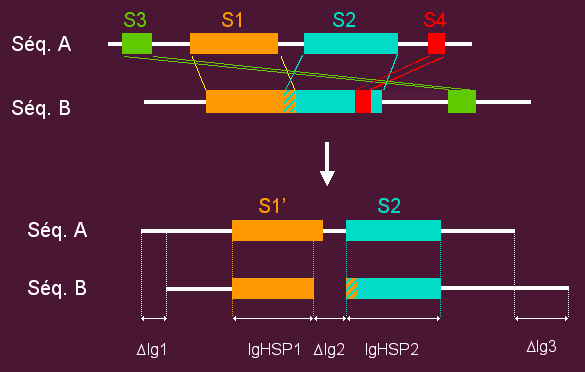
- For each pair of sequences, Homologous Segment Pairs (HSPs) that are not compatible with a global alignment are removed (see example).
- Two sequences in a pair are included in the same family if:
- The remaining HSPs cover at least 80% of the proteins length.
- Their similarity is greater or equal to 50% (two amino-acids are considered similar if the BLOSUM62 similarity score is positive).
- Both sequences are complete.
- We use simple transitive links to build our families. If a pair of sequences named A + B and a pair of sequences B + C fulfill the conditions listed above, then A, B and C are integrated in the same family, this even if the pair A + C does not fulfill these conditions.
- Once families of complete protein sequences have been build, partial sequences (longer than 100 AA or at least 50% of the length of the complete proteins) are included in the classification. A partial sequence matching with a complete protein is included in its family if:
- The remaining HSPs cover at least 80% of the partial protein length.
- Their similarity is greater or equal to 50%.
- Short partial sequences (less than 100 AA and less than 50% of the length of the complete proteins) are not included in the classification.
{kind=link}
Alignments
For each family, protein sequences are aligned using CLUSTALW 1.7. All the default parameters are used excepted that the "Fast/Approximate" option is preferred for pairwise alignments.
Phylogenetic trees
The distance used to build the phylogenetic trees is the observed divergence. When the distance matrix is complete, phylogenetic trees are computed with BIONJ. When the matrix is uncomplete (i.e. when there are partial sequences in the family that don't overlap with each others), we use the Triangle method (Alain Guenoche, unpublished).